

Исходя из вышеперечисленного в предыдущей статье рациона питания озёрных цихлид в естественной среде обитания, они подразделяются на несколько основных типов, а именно:
— питающиеся листьями и стеблями высших растений;
— питающиеся фитопланктоном (планктоном растительного происхождения);
— водорослеядные или альгофаги (Algophages), т.е. питающиеся водорослями и микроживотными, живущими в них, но, где основную долю питания составляет всё же пища растительного происхождения;
— водорослеядные, питающиеся водорослями (с живущими в них микроживотными), покрывающими листья высших растений (перифитоном).
— питающиеся себе подобными меньшего размера (молодью цихлид или другими рыбами некрупных видов, например, озёрными сардинами) — рыбоядные хищники (Piscivore);
— питающиеся икрой или личинками себе подобных — паедофаги (Paedophage);
— питающиеся плавниками или чешуёй себе подобных — лепидофаги (Lepidophage);
— питающиеся несъеденными остатками пищи себе подобных;
— питающиеся микроживотными (зоопланктоном и, отчасти, фитопланктоном), обитающими в открытых водах — планктофаги (Planctophage);
— питающиеся беспозвоночными (личинками насекомых и ракооборазными), живущими в донном грунте или в непосредственной близости от него — бентофаги (Benthivore или Benthophage);
— питающиеся моллюсками — моллюскоядные (Molluscivore).
— питающиеся органическими и микроорганическими остатками пищи растительного и животного происхождения;
— питающиеся выделениями других рыб (фикалиями), которые содержат непереваренные остатки пищи растительного и (или) животного происхождения.
— питающиеся осадочными отложениями и органическим «мусором», состоящим из погибших микроорганизмов, остатков жизнедеятельности рыб и животных и песчанной взвеси, а также живущими в детрите микроживотными.
Очень часто, даже не зная к какому из данных выше типов относится та или иная цихлида, по форме её корпуса и головы, по форме и размеру рта, по форме и строению зубов можно сделать предварительный вывод о кормовой специализации (амплуа) и промысловых особенностях того или иного вида, о его «охотничьих» повадках.
По приведённым ниже фотографиям, показывающим форму передней части тела, а также форму головы и рта, можно с той или иной долей вероятности судить о принадлежности некоторых видов цихлид к тому или иному типу, о технике и технологии питания, а также об их кормовой специализации.
Рыбоядные хищники:
 |
— Aristochromis christyi |
|---|---|
 |
— Champsochromis caeruleus |
 |
— Dimidiochromis compressiceps |
 |
— Nimbochromis livingstoni |
 |
— представитель рода Ramphochromis |
Планктофаги:
 |
— Copadichromis mloto |
|---|---|
 |
— Copadichromis mloto |
Альгофаги:
 |
— Labeotropheus trewavasae |
|---|---|
 |
— Petrotilapia tridentiger |
 |
— представитель видового комплекса Pseudotropheus zebra |
 |
— Pseudotropheus acei |
 |
— представитель рода Tropheops |
Паедофаги:
 |
— Caprichromis orthognathus |
|---|---|
 |
— Diplotaxodon greenwoodi |
 |
— Naevochromis chrysogaster |
 |
— Protomelas spilopterus |
Лепидофаги:
 |
— Corematodus shiranus |
|---|---|
 |
— Corematodus taeniatus |
 |
— Docimodus johnstoni |
 |
— Geniochromis mento |
 |
— Melanochromis lepidiodaptes |
Толстогубые (извлекающие пищу из расщелин, щелей, ямок и углублений):
 |
— Chilotilapia euchilus |
|---|---|
 |
— Melanochromis labrosus |
 |
— Placidochromis milomo |
Сборщики перифитона (микроживотных и водорослей с листьев высших растений):
 |
— Cyathochromis obliquidens |
|---|---|
 |
— Hemitilapia oxyrhynchus |
Чистильщики (эктопаразитов с кожного покрова рыб):
 |
— Metriaclima (Pseudotropheus) crabro |
|---|
Моллюскоядные:
 |
— Trematocranus placodon |
|---|
А по этим фотографиям, показывающим форму рта и ротовой полости, а также форму, строение и расположение зубов можно предварительно судить о кормовом амплуа той или иной цихлид:
 |
— форма рта и ротовой полости, а также расположение острых, частых зубов рыбоядного хищника из рода Bathybates |
|---|---|
 |
— форма рта и «прикус» представителя рода Bathybates |
 |
— форма и строение зубов представителя рода Cyathopharynx |
 |
— форма рта и расположение зубов детритофага из рода Cyathopharynx |
 |
— форма рта и расположение зубов представителя рода Spathodus |
 |
— форма рта и расположение зубов представителя рода Trematocara |
 |
— форма рта и расположение мелких, многорядных зубов-тёрок Petrochromis polyodon |
 |
— форма рта и головы рыбоядного хищника Altolamprologus compressiceps |
 |
— форма рта и строение ротового аппарата планктофага из рода Cyprichromis |
 |
— форма рта и расположение крепких, загнутых внутрь зубов типичного лепидофага Perissodus straeleni |
 |
— форма рта с толстыми губами представителя рода Placidochromis |
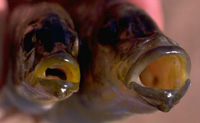 |
— форма рта и ротовой полости Naevochromis chrysogaster |
 |
— строение и расположение глоточных зубов типичного рыбоядного хищника |
Попробуем рассмотреть кормовую специализацию и технологию питания, объекты питания и средства их обнаружения и извлечения, а также технику, используемую цихлидами для выполнения важнейшей составляющей жизнеобитания — питания — с помощью приведённой ниже таблицы:
| Растительноядные | Высшие растения | Отрывание листьев и стеблей или отдельных их частей при помощи зубов (захват) и корпуса (движения в сторону и назад) | В 3 раза больше общей длины тела | Protomelas similis (М) |
|---|---|---|---|---|
| Растительноядные | Фитопланктон | Перемалывание клеток фитопланктона при помощи множества коротких зубов, расположенных на глоточной кости | В 5 раз больше общей длины тела | Copadichromis pleurostigma (М); Copadichromis trimaculatus (М) |
| Водорослеядные | Одноклеточные водоросли (’кремнезёмки’) и крупинки песка | Состригание одноклеточных водорослей (рыхлый материал) с водорослевого каркаса обрастаний при помощи множества тонких, подвижных зубов-’трезубцев’ и рта с толстыми губами, которыми герметизируется место извлечения пищи | В 10 раз больше общей длины тела (длина желудочно-кишечного тракта взрослых особей достигает 2-х метров) | Petrochromis (Т) |
| Водорослеядные | Илистые водоросли | Отрывание ила от осадочных отложений, зажав его зубами и двигая корпусом назад | В 2,5 раза больше общей длины тела | Triglachromis otostigma (Т) |
| Водорослеядные — сборщики перифитона (Periphyton) | Водоросли и микроживотные | Соскабливание водорослей и микроорганизмов с листьев высших растений, плотно захватив их ртом и двигая корпусом назад, повторяя процедуру 3-4 раза | В 1,5-2 раза больше общей длины тела | Hemitilapia oxyrhynchus (М); Cyathochromis obliquidens (M) |
| Водорослеядные | Водоросли и микроживотные | Соскребание или общипывание водорослей с субстрата под углом 45 гр. (и менее) при помощи низкорасположенного рта и зубов-’скребков’ | В 1,5-2 раза больше общей длины тела | Labeotropheus (М); Tropheops (М); Ps. elongatus-Komplex (М); Cynotilapia (М) |
| Водорослеядные | Водоросли и микроживотные | Соскребание водорослей с субстрата или расчёсывание водорослевых грядок под углом 90 гр. при помощи пряморасположенного рта и зубов-’расчёсок’ | В 1,5-2 раза больше общей длины тела | Petrotilapia (М); Metriaclima (М); Pseudotropheus (М; крупные виды); Melanochromis (М) |
| Водорослеядные | Водоросли и микроживотные | Грабление водорослевых грядок редкорасположенными зубами | В 1,5-3 раза больше общей длины тела | Eretmodus (Т); Spathodus (Т); Tanganicodus (Т) |
| Водорослеядные | Водоросли и микроживотные | Откусывание или отрывание водорослей от субстрата при помощи острых, плотнорасположенных зубов и головы (рывок в сторону) | В 1,5-2 раза больше общей длины тела | Pseudotropheus demasoni (М); Melanochromis maingano (M); Tropheus (Т) |
| Водорослеядные | Водоросли и микроживотные | Высасывание содержимого углублений шпата или шероховатостей скал при помощи толстых, мясистых губ (пневмо-гидравлический замок) и ротовой полости (создание вакуума) | В 1,5-2 раза больше общей длины тела | Placidochromis milomo (М); Chilotilapia euchilus (М); Protomelas taeniolatus (М) |
| Водорослеядные | Водоросли и микроживотные | Выклёвывание содержимого углублений шпата при помощи узкого, удлинённого рта и острых зубов | В 1,5-2 раза больше общей длины тела | Labidochromis (М); Pseudotropheus (М; мелкие виды) |
| Водорослеядные | Водоросли и микроживотные | Грабление илистого слоя, лежащего на песке, и извлечение илистых водорослей и микроживотного материала | В 4 раза больше общей длины тела | Otopharynx heterodon (М); Otopharynx spec. ’aeromarginatus’ (М) |
| Плотоядные — рыбоядные хищники | Молодь цихлид или взрослые особи мелких видов рыб группы ’Usipa’ (озёрные сардины) | Обнаружение добычи при помощи чувствительных органов, захват и заглатывание её с использованием удлинённой головы, острых зубов и вместительной ротовой полости | 60-90% от общей длины тела | Dimidoichromis (М); Champsochromis (М); Tyranochromis (М); Nimbochromis (М); Taeniochromis (М); Hemitaeniochromis (М); Stigmatochromis (М); Exochochromis anafenys (М); Aristochromis christyi (М); Placidochromis johnstoni (М); Lepidiolamprologus (Т); Bathybates (Т); Haplotaxodon (Т); Boulengerochromis (Т) |
| Плотоядные — рыбоядные хищники | Молодь рыб | Извлечение добычи из узких расщелин (иногда с поворотом тела на 90 гр.) при помощи сплюснутого по бокам корпуса, удлинённой морды и очень прочной чешуи | 60-90% от общей длины тела | Altolamprologus (T); |
| Плотоядные — ’паедофаги’ | Молодь рыб | Извлечение добычи из узких расщелин (иногда с поворотом тела на 90 гр.) при помощи сплюснутого по бокам корпуса, удлинённой морды и очень прочной чешуи | 60-90% от общей длины тела | Telmatochromis (Т) |
| Плотоядные — ’паедофаги’ | Икра и личинки рыб | Выкрадывание добычи c использованием фактора внезапности и многочисленности (рассеивание внимания) | 60-90% от общей длины тела | Caprichromis liemi (М); Caprichromis orthognatus (M); Diplotaxodon greenwoodi (M); Naevochromis chrysogaster (M); Protomelas spilopterus (M); Hemitaeniochromis spec. ’paedophage’ (М) |
| Плотоядные — ’лепидофаги’ | Чешуя рыб | Выбивание, соскребание или откусывание чешуи от корпуса других видов цихлид при помощи крепких челюстей и зубов и её заглатывание | 60-90% от общей длины тела | Docimodus (М); Corematodus (М); Genyochromis mento (М; доп. источник питания); Perissodus (Т); Plecodus (Т); Melanochromis lepidiodaptes (M) |
| Плотоядные | Плавники рыб | Обгрызание плавников других цихлид при помощи мелких, острых зубов | 60-90% от общей длины тела | Genyochromis mento (М); Docimodus johnstoni (M) |
| Плотоядные — ’сборщики мусора’ | Органические и микроорганические отходы | Подъедание несъеденных остатков пищи от ’трапезы’ крупных рыб, а также фикалий растительноядных видов, которыми богаты осадочные отложения | В 1-1,5 раза больше общей длины тела | Ctenopharynx pictus (М); Otopharynx lithobates (М); Otopharynx walteri (М); Otopharynx spec. ’cave’ (М) |
| Плотоядные — ’озёрные санитары’ | Несъеденные остатки пищи | Подбирание остатков пищи, выходящих через жабры крупных цихлид (Taeniolethrinops praeorbitalis) вместе с крупинками песка, с использованием удобной позиции (сзади ’хозяина’ и ниже по глубине), а также поиск съедобных частиц во взрыхлённом ’хозяином’ грунте. | В 1-1,5 раза больше общей длины тела | Placidochromis phenochilus (М); Cyrtocara moorii (М); Otopharynx selenurus (М); Protomelas annectens (М); Placidochromis electra (М) -последние два вида могут пользоваться и другими источниками пропитания. |
| Плотоядные — ’планктофаги’ | Зоопланктон и, частично, фитопланктон | Захват одиночных или групповых объектов при помощи рта-’ловушки’ или ’выдвигающегося’ рта и вместительной ротовой полости | В 1,5- 2 раза больше общей длины тела | Cynotilapia (М); все виды группы ’Utaka’ (М); Cyprichromis (Т); Paracyprichromis (Т) |
| Плотоядные — «бентофаги’ | Личинки насекомых и моллюски | Обнаружение добычи при помощи чувствительных сенсорных органов, погружение головы в песок и захват добычи ртом с последующим отфильтровыванием песка через жабры | В 1-1,5 раза больше общей длины тела | Aulonocara (М); Trematocranus (М); Alticorpus (М); Xenotilapia (Т); Trematocara (Т); Aulonocranus (Т); Telotrematocara (Т) |
| Плотоядные — ’бентофаги’ | Личинки насекомых и моллюски | Всасывание ртом добычи вместе с песком и отфильтровывание последнего через жабр | 1-1,5 раза больше общей длины тела | Gnatochromis permaxillaris (Т) |
| Плотоядные — ’бентофаги’ | Личинки насекомых и моллюски | Всасывание и выброс ртом струи воды в рыхлый грунт (в место обнаружения объекта) с целью размывания этого места и захвата ’оголившейся’ добычи | В 1,5-2 раза больше общей длины тела | Protomelas fenestratus (М; многие геоварианты) |
| Плотоядные — ’бентофаги’ | Личинки насекомых и моллюски | Переворачивание или опрокидывание небольших камней при помощи крепких челюстей и захват, обнаруженной под камнем, добычи | В 1,5-2 раза больше общей длины тела | Mylochromis labidodon (М); Placidochromis spec. ’johnstoni solo’ (М) |
| Плотоядные | Личинки насекомых и ракообразные | Собирание добычи из шероховатостей скал (пневмо-гидравлический замок), даже без создания вакуума, при помощи толстых, гибких, шероховатых губ, чётко повторяющих контур скал | В 1,5-2 раза больше общей длины тела | Lobochilotes labiatus (Т) |
| Плотоядные | Личинки насекомых | Извлечение (высасывание) добычи из узких трещин и щелей при помощи утолщённых губ и удлинённых головы и рта | В 1,5-2 раза больше общей длины тела | Protomelas ornatus (М); Lichnochromis aciticeps (М) |
| Плотоядные — ’бентофаги’ | Молодь рыб, креветки, мелкие рачки | Вычерпывание добычи из заиленного грунта с использованием рта, как черпака (ковша) | Приблизительно 90% от общей длины тела | Reganochromis calliurus (Т) |
| Плотоядные — моллюскоядные хищники | Большие беспозвоночные и крупные ракообразные (крабы) | Нападение на добычу и разгрызание прочного панциря при помощи сильных фарингиальных костей обоих челюстей с последующим поглощением только плоти | В 1-1,2 раза больше общей длины тела | Mylochromis epichorialis (М); Limnochromis (Т) |
| Плотоядные | Ракообразные и улитки | Склёвывание добычи с листьев, стеблей и корней высших растений , а, иногда, и с поверхности грунта, окружающего растения | В 1-1,5 раза больше общей длины тела | Protomelas kirkii (М); Protomelas labridens (М); Protomelas marginatus (М); Protomelas pleurotaenia (М) |
| Плотоядные — ’моллюскоядные’ | Моллюски (улитки) | Раскалывание панциря улитки при помощи глоточных зубов и высасывание содержимого | В 1,5-2 раза больше общей длины тела | Chilotilapia rhoadesi (М); Mylochromis melanotaenia (М); Mylochromis balteatus (М) |
| Плотоядные — ’чистильщики’ | Кожные паразиты (эктопаразиты) | Склёвывание с гладкой кожи крупных озёрных сомов, паразитирующих на них рыбьих вшей (Argulus africanus) | В 1,2-1,5 раза больше общей длины тела | Pseudotropheus crabro (М) |
| Всеядные | Пища животного и растительного происхождения | Поедание любых съедобных, движущихся или неподвижных объектов | В 1-1,2 раза больше общей длины тела | Astatoreochromis (Т); Astatotilapia (Т); Jodotropheus sprengerae (М) |
| Детритоядные | Детрит и рыхлый материал обрастаний (одноклеточные водоросли) | Захват съедобных объектов вместе с детритом или только детрита | В 4-5 раз больше общей длины тела | Cyathopharynx (Т); Ophthalmotilapia (Т) |
В последнем столбце таблицы буквами «М» и «Т» обозначена принадлежность рода или вида к соответствующему озеру (Малави или Танганьика).
В приведённой таблице показаны наиболее типичные варианты кормовой специализации, техники и технологии питания, хотя не исключены и нетипичные варианты, к примеру такие как , выбивание и поедание глаз и т.п.
В таблице также приведены соотношения длины кишечника цихлиды к общей длине её тела (относительная длина кишечника), из чего можно сделать вывод, что цихлиды, питающиеся трудноперевариваемой пищей (растения, водоросли, детрит), имеют более длинный кишечник, у всеядных видов — он более короткий, но всё ещё длиннее общей длины тела, а у плотоядных видов, особенно у рыбоядных хищников, — кишечник, как правило, соизмерим с длинной тела или немного короче. Если судить о толщине стенок кишечника и желудка, то у цихлид, питающихся крупной добычей (рыбоядные хищники), они более толстые, а у цихлид, питающихся микроживотными и водорослями, — тонкие.
Многие водорослеядные виды цихлид, приведённые в таблице, можно условно отнести к всеядным видам, т.к. их рацион питания составляет пища животного и растительного происхождения, но значительную долю рациона всё же составляет растительная составляющая, поэтому и вопрос этот можно пока оставить открытым до тех пор, пока исследователи абсолютно точно не разберутся с рационом и кормовой специализацией.
Тоже самое можно сказать и о плотоядных видах ( исключая рыбоядных хищников), незначительную долю рациона которых составляет растительная часть и кишечник которых всё же длиннее, чем у чисто плотоядных видов.
Ниже приведена относительная длина кишечника некоторых видов танганьикских цихлид (первая цифра — длина кишечника, вторая цифра — общая длина тела):
Bathybates fasciatus — 0,5 : 1
Bathybates ferox — 0,5 : 1
Bathybates leo — 0,7 : 1
Bathybates minor — 0,6 : 1
Boulengerochromis microlepis — 1,8 : 1
Cardiopharynx schoutedeni — 3,3 : 1
Cunningtonia longiventralis — 5 : 1
Cyathopharynx foai — 3 : 1
Cyathopharynx furcifer — 3 : 1
Cyphotilapia frontosa — 0,8 : 1
Cyprichromis microlepidotus — 0,8 : 1
Ectodus descampsi — 0,8 : 1
Eretmodus cyanostictus — 2,5 : 1
Gnathochromis permaxillaris — 1,3 : 1
Haplochromis burtoni — 3,3 : 1
Lepidiolamprologus elongatus — 0,4 : 1
Lestradea perspicax perspicax — 3,5 : 1
Lestradea perspicax stappersi — 2,8 : 1
Limnochromis auritus — 0,3 : 1
Limnochromis abeelei — 0,6 : 1
Limnochromis staneri — 1 : 1
Limnotilapia dardennii — 1,8 : 1
Neolamprologus brevis — 0,8 : 1
Neolamprologus brichardi — 0,6 : 1
Neolamprologus furcifer — 0,5 : 1
Neolamprologus kungweensis — 0,4 : 1
Neolamprologus savoryi — 0,7 : 1
Neolamprologus signatus — 0,4 : 1
Neolamprologus toae — 0,6 : 1
Oreochromis niloticus — 6,2 : 1
Oreochromis tanganicae — 5,6 : 1
Petrochromis orthognatus — 6,4 : 1
Petrochromis trewavasae — 10 : 1
Pseudosimichromis curvifrons — 3,0 : 1
Simochromis babaulti — 3,1 : 1
Simochromis diagramma — 3,2 : 1
Simochromis marginatus — 5,1 : 1
Spathodus erythrodon — 2,5 : 1
Tangachromis dhanisi — 1 : 1
Tanganicodus irsacae — 1,75 : 1
Telmatochromis bifrenatus — 0,6 : 1
Telmatochromis dhonti — 0,9 : 1
Telmatochromis temporalis — 1,6 : 1
Telmatochromis vittatus — 1,1 : 1
Trematocara caparti — 0,5 : 1
Trematocara kufferathi — 0,4 : 1
Trematocara marginatum — 0,6 : 1
Trematocara nigrifrons — 0,5 : 1
Trematocara stigmaticum — 0,4 : 1
Trematocara unimaculatum — 0,6 : 1
Trematocara variabile — 0,4 : 1
Triglachromis otostigma — 2,4 : 1
Tropheus Moorii — 4,8 : 1
Tylochromis polylepis — 1,6 : 1
Xenotilapia bathyphilus — 0,8 : 1
Xenotilapia boulengeri — 0,7 : 1
Xenotilapia burtoni — 1 : 1
Xenotilapia caudafasciata — 0,8 : 1
Xenotilapia longispinis — 1 : 1
Xenotilapia nigrolabiata — 1,2 : 1
Xenotilapia ochrogenys — 0,8 : 1
Xenotilapia ornatipinnis — 0,7 : 1
Xenotilapia papilio — 0,9 : 1
Xenotilapia sima — 0,7 : 1
Технику питания некоторых видов малавийских и танганьикских цихлид можно увидеть на приведённых ниже фотографиях:
а) «прослушивание» дна с помощью сенсорных органов самкой Aulonocara:

б) зарывание головы в песчаный грунт для извлечения добычи представителем рода Mylochromis:

в) зарывание головы в песчаный грунт для извлечения добычи представителем рода Xenotilapia:

г) отрывание обрастаний от скал при расположении тела и рта под углом 45 градусов к субстрату:

д) склёвывание добычи с рыхлого субстрата под углом 45 градусов:

е) извлечение добычи из расщелины с поворотом корпуса на 90 градусов:

ж) отрывание обрастаний от скал при расположении тела и рта под углом 90 градусов к субстрату:

з) соскабливание водорослей и детрита со скального субстрата самцом Ophthalmotilapia при расположении тела и рта под углом 90 градусов к объекту:

Для того, чтобы более полно представить себе стратегию и тактику рыбоядных хищников по поиску, обнаружению и захвату потенциальных жертв, а также технические приёмы, используемые другими типами цихлид для решения вопроса пропитания, обратимся к нижеприведённым рисункам, которые с определённой долей наглядности демонстрируют тактические варианты решения этой жизненноважной для цихлид дилемы:
1. Цихлиды, питающиеся донным и придонным кормом (Benthivores):
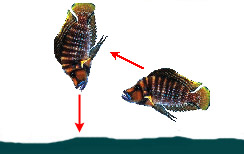 |
— Осуществляет поиск при плавании параллельно субстрату; при обнаружении добычи занимает позицию перпендикулярно дну и атакует | ||
|---|---|---|---|
 |
— Осуществляет поиск и обнаружение добычи, при нахождении на большой дистанции | ||
 |
— Осуществляет поиск и обнаружение добычи, при нахождении на средней дистанции | ||
 |
— Осуществляет поиск и обнаружение добычи, находясь в непосредственной близости от неё | ||
 |
— Осуществляет поиск и обнаружение добычи в донных или придонных углублениях (ямках, отверстиях) субстрата | ||
 |
— Осуществляет поиск и обнаружение добычи в расщелинах (трещинах) скального субстрата | ||
 |
— Осуществляет поиск и обнаружение добычи при помощи хвостового плавника, разбрасывая им рыхлый донный субстрат | ||
 |
— Осуществляет поиск и обнаружение добычи, зарывая нижнюю часть корпуса в рыхлый песчанный грунт до уровня рта при помощи брюшных и анального плавников | ||
 |
— Осуществляет поиск и обнаружение добычи путём зарывания рта и части головы в рыхлый песчанный грунт | ||
 |
— Осуществляет поиск и обнаружение добычи непосредственно на поверхности донного субстрата |
 |
— Соскабливает (соскребает) водоросли или детрит с донного или скального субстрата, располагаясь под углом 30-45 гр. к нему (низкорасположенный рот) | ||
|---|---|---|---|
 |
— Отрывает (отдирает) водоросли и детрит от скального субстрата, располагаясь перпендикулярно (или почти перпендикулярно) субстрату (пряморасположенный рот) | ||
 |
— Соскабливает (соскребает) водоросли и детрит со скального субстрата, располагаясь под углом 30-45 гр. к нему (низкорасположенный рот) | ||
 |
— Отрывает (отдирает) водоросли от небольших, отдельнолежащих глыб и камней, располагаясь почти перпендикулярно к объекту питания (пряморасположенный рот) | ||
 |
— Соскабливает (соскребает) водорослевой налёт (перифитон) с листьев и стеблей высших растений (пряморасположенный рот) | ||
 |
— Собирает водоросли, детрит и крупинки песка со скального субстра та, располагаясь параллельно к нему (низкопасположенный рот) |
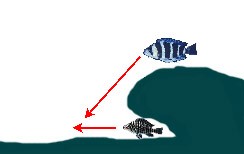 |
— Нападение из засады, расположенной на нависающей скале, на выплывающую из убежища цихлиду | ||
|---|---|---|---|
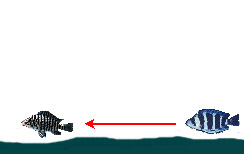 |
— Преследование добычи на открытом участке донной поверхности | ||
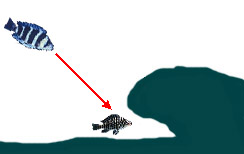 |
— Преследование уходящей в убежище добычи из позиции выше по глубине | ||
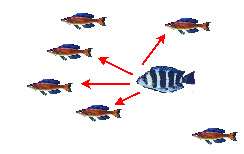 |
— Нападение на цихлид, находящихся в составе стаи | ||
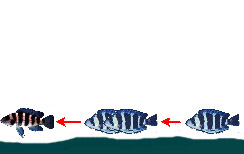 |
— Преследование и нападение на добычу ’из-за спины’ других цихлид |
 |
— Ловля (захват) одиночных или групповых объектов в открытой воде (зона открытых вод) | ||
|---|---|---|---|
 |
— Ловля (захват) одиночных или групповых объектов на удалении от донного и скального субстратов (зона открытых вод над твёрдым субстратом) | ||
 |
— Ловля (захват), преимущественно, одиночных объектов в водяном столбе вблизи своего убежища-нерестилища (зона открытых вод над песчанным субстратом) | ||
 |
— Ловля (захват) одиночных или групповых объектов вблизи скального субстрата (зона открытых вод над скальным субстратом) |
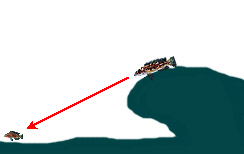 |
— Обнаружение добычи из засады, расположенной на скале выше по глубине (место наилучшего обзора) … | ||
|---|---|---|---|
 |
… и внезапное нападение на жертву на открытом участке дна | ||
 |
— Обнаружение добычи из засады, расположенной под нависающей скалой, и внезапное нападение на жертву на открытом участке дна | ||
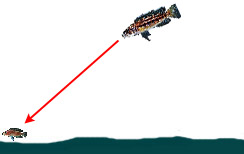 |
— Обнаружение и нападение на добычу, находящуюся на открытом участке дна, из позиции выше по глубине (охота в режиме «рейдера» или «одинокого волка») | ||
 |
— Обнаружение и нападение на добычу, находящуюся на открытом участке дна, под прикрытием другой нехищной цихлиды из позиции выше по глубине | ||
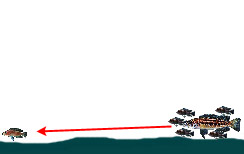 |
— Обнаружение и нападение на добычу на открытом участке донной поверхности с использованием прикрытия стаи нехищных цихлид | ||
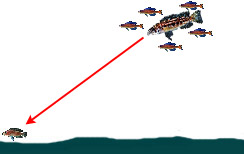 |
— Обнаружение и нападение на добычу, находящуюся на открытом участке дна, с использованием позиции выше по глубине и под прикрытием стаи нехищных цихлид | ||
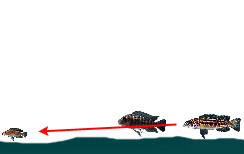 |
— Обнаружение и нападение на добычу на открытом участке дна с использованием позиции «из-за спины» другой, нехищной цихлиды | ||
 |
— Ночная охота представителя рода Cyphotilapia на своих потенциальных жертв из рода Cyprichromis с использованием преимущественной позиции по глубине, тёмного времени суток и «полусонного» состояния добычи (или её ослабленного состояния) |
| Вода | Белок | Жир | |
|---|---|---|---|
| Циклоп | 80% | 9% | 2-3% |
| Дафния | 95% | 2,5% | 0,5% |
| Мотыль | 88% | 6,5% | 2% |
| Кулекс | 82% | 10% | 4% |
Из приведённой таблицы можно сделать однозначный вывод, что из ракообразных (циклоп и дафния) основным источником протеина (белка) в кормовом спектре является циклоп, а из беспозвоночных (личинок насекомых), коими являются мотыль и кулекс, — кулекс. Ввиду многообразия видов циклопа в озёрах и его многочисленности он по праву считается основой кормового рациона животного происхождения для многих представителей семейства цихлид, даже для многих крупных видов, составляющих группу ’Utaka’, и не только — наглядный пример, Fossorochromis rostratus, который в природе отдаёт предпочтение циклопу и только в ’голодное время’ или в нерестовый период кормится вблизи дна другими беспозвоночными.
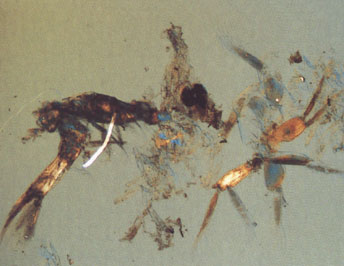
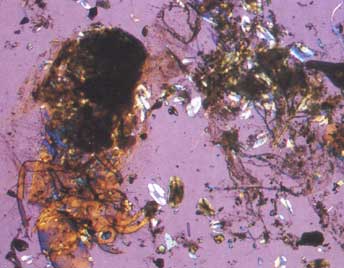
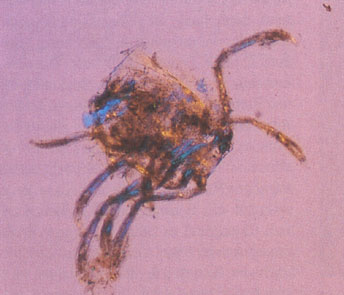
Наглядно увидеть, что представляет из себя содержимое кишечника и желудка различных озёрных цихлид, можно на снимках, сделанных при исследовании пищеварительного тракта рыб под микроскопом, которые предлагаются вниманию цихлидников:
 |
— содержимое желудка, основную часть которого занимает циклоп и крупная личинка мухи-однодневки; |
|---|---|
 |
— содержимое кишечника Neolamprologus furcifer, состоящее их фрагментов личинок насекомых; |
 |
— содержимое кишечника Tanganicodus irsacae, состоящее из фрагментов личинок насекомых, ракушкообразных, водорослей и частичек песка; |
 |
— фрагмент водяного клеща из кишечника Neolamprologus furcifer; |
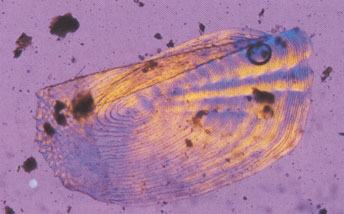 |
— непереваренная ещё рыбья чешуйка из желудка Perissodus microlepis; |
 |
— содержимое кишечника Tropheus moorii, состоящее из фрагментов водорослей- кремнезёмок и множества частичек песка |
В зависимости от времени года, биотопа обитания и локальных катаклизмов питание цихлид в озёрах может быть относительно обильным и весьма скудным («голодное время»). Это оказывает влияние, как на жизнеобитание цихлид, так и на манеру их повседневного поведения. Но, оказывается, на рацион питания влияют не только вышеперечисленные факторы — многие виды цихлид могут радикально изменять рацион питания в течении дня. Например, ранним утром, когда ещё достаточно темно, они поедают трудноперевариваемые или неперевариваемые вовсе зелёные водоросли (Anabaena), что для них, как правило, нетипично. Почему именно эти водоросли и почему ранним утром? Ведь зелёные водоросли практически не перевариваются в средней кишке? А дело обстоит так: ранним-ранним утром кислотность (pH) желудочного сока ещё достаточно высока (после ночного отдыха лишь к рассвету кислотность достигает своего низшего значения), что позволяет делать стенки клеток водорослей более проницаемыми. Участок средней кишки с непереваренными зелёными водорослями, сохранившими в ней свой природный цвет, уже во второй половине дня или к вечеру заполняется новыми порциями корма, но другого содержания — это циклоп и личинки насекомых. К этому времени зелёные водоросли под воздействием желудочного сока приобретают коричневый цвет, а недавно съеденные циклоп и личинки насекомых уже нормально перевариваются в заднем, кишечном участке. Остаётся открытым вопрос — почему цихлиды, как правило, не питающиеся зелёными водорослями, всё же поедают их ранним утром? Исследователи озёр полагают, что такое несвойственное поедание служит сигналом к «включению» желудочных органов, вырабатывающих кислотный секрет (органы секреции), активно участвующий в пищеварительном процессе. Такую же функцию отчасти выполняют и детрит, и песок, и частички известнякового шпата.
Поиск объекта питания (добычи) у различных видов цихлид осуществляется по разному, как в одиночку, так и стайно. Рыбоядные хищники могут охотиться или в режиме ’одинокого волка’ или в режиме ’волчьей стаи’; планктоноядные — как правило, группируются в большие стаи, как одновидовые, так и смешанные, как однополые, так и разнополые (с присутствием в стае самок незрелых самцов); бентофаги и альгофаги — как правило, собираются в небольшие по количеству стаи (самки), а территориальные самцы питаются исключительно в одиночном режиме.
На приведённых ниже фотографиях можно наглядно увидеть некоторые поисковые группы, часто называемые «фуражными»:
а) небольшая поисковая группа Cyathopharynx, состоящая из трёх незрелых самцов:

б) небольшая смешанная поисковая группа самок малавийских цихлид (поиск на песчаном грунте):

в) небольшая одновидовая поисковая группа самок малавийских цихлид:

г) большая поисковая группа молодых особей Fossorochromis rostratus (поиск на песчанном участке дна):

д) большая поисковая группа Cyprichromis в открытых водах:

е) большая поисковая группа Tropheus brichardi «Mpimbwe» и рядом с ней чуть выше по глубине небольшая стайка Cyathopharynx:

Процесс питания озёрных цихлид можно условно разделить на несколько временных этапов, уяснение сути которых позволит аквариумистам-цихлидникам осознанно и правильно подойти к вопросу кормления своих любимцев при аквариумном содержании:
— поиск объекта питания при помощи зрительных или других чувствительных органов;
— обнаружение объекта питания и его непродолжительное изучение;
— извлечение объекта питания из субстрата, или его отделение от субстрата (отрывание, соскребание, соскабливание, — — —
— откусывание, высасывание и т.п.), или захват добычи (мелкой рыбы, зоопланктона) в толще воды;
— отфильтровывание съедобных составляющих корма от несъедобных;
— заглатывание объекта питания целиком или его перетирание (разделение) при помощи глоточных зубов;
— переваривание пищи в желудке и кишечнике;
— выведение переваренной пищи из организма.
Процесс «добывания хлеба насущного» для цихлид, обитающих в естественной среде обитания, сопряжён с целым рядом неизбежных трудностей (сезонные явления, жесточайшая конкуренция, опасность со стороны всевозможных хищников и т.д.), которые им приходится преодолевать, затрачивая много усилий и времени. Поэтому процесс питания и его этапы не скоротечен и протекает относительно медленно, но неумолимо. Не всегда найденный и съеденный объект питания может компенсировать энергетические затраты, потраченные на его поиск, обнаружение и извлечение, а если учесть другие энергозатраты, как то: борьба за нерестовые или кормовые угодья, охрана гнездовья и потомства от посягательств, строительство родового гнезда, вынашивание икры и эмбрионов во рту и т.д. и т.п., то создаётся впечатление, что цихлиды большую часть времени борются за своё существование и занимаются вопросами воспроизводства, и лишь ничтожная часть времени уходит у них на то, чтобы хоть частично насытиться. У них просто нет ни времени, ни возможности наедаться «до отвала», да и от сверхсытой, ожиревшей рыбы мало толку — она уже не производитель, не любовник, не боец ! И матушка-природа очень грамотно регулирует этот процесс !
Я думаю, что с помощью этой и предыдущей статьи любители получили определённый набор информации о рационе питания и кормовой специализации, технике и технологии питания озёрных цихлид в природе. Настал черёд более детально рассмотреть вопрос кормления в условиях домашнего содержания ’пёстрых окуней’ — этим мы и займёмся в следующей статье цикла статей о питании и кормлении.
Автор статьи выражает искреннюю благодарность доктору Михаэлю Оливеру (Dr. Michael K. Oliver — www.malawicichlids.com) за любезное разрешение использовать некоторые фотоматериалы его сайта для иллюстрирования данной статьи. В статье также использованы графические иллюстрации (рисунки) с сайта французского аквариумиста-исследователя Эрика Женевиля (Eric Genevelle — www.tanganyika-cichlids.com).